
Barwniki fotosyntetyczne
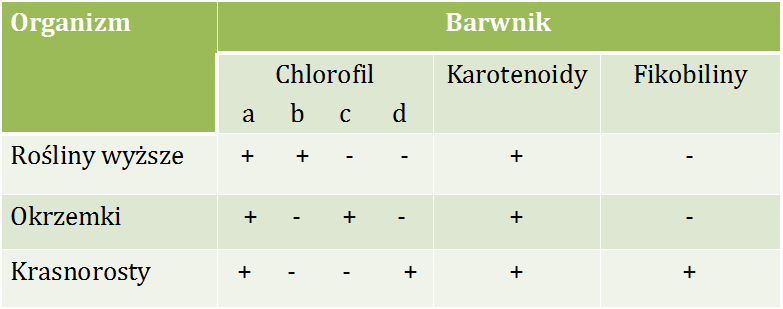
Barwniki fotosyntetyczne występują w aparatach fotosyntetycznych fotoautotrofów. Wyróżnia się trzy klasy barwników fotosyntetycznych: chlorofile, karotenoidy oraz fikobiliny. Występują one u różnych organizmów (Tab.4.1) [14, 15].
Tabela 4.1. Występowanie barwników fotosyntetycznych w wybranych grupach organizmów [14,15]
Chlorofile
Chlorofile stanowią główną grupę barwników fotosyntetycznych. Występują powszechnie u roślin zielonych i glonów, nadając im zielone zabarwienie. Absorbują one światło w zakresie widzialnym przy długościach fali 420 – 460 nm (światło niebieskie) oraz 650 – 700 nm (światło czerwone) [14].
W wyniku absorpcji promieniowania cząsteczka chlorofilu ulega wzbudzeniu (światło niebieskie – drugi stan wzbudzony; światło czerwone – pierwszy stan wzbudzony). W związku z niską trwałością wzbudzonego chlorofilu, zachodzi szybki powrót elektronów do stanów podstawowych, co wiąże się z uwolnieniem energii. Proces ten może przebiegać na trzy sposoby:
poprzez przekazanie energii wzbudzenia elektronów do cząsteczki akceptorowej;
poprzez rozproszenie termiczne;
poprzez emisję energii w postaci niżej energetycznego fotonu (fluorescencja – w chlorofilach związana jest ona z pierwszym stanem wzbudzenia) [14].
Wyróżnia się kilka rodzajów chlorofili (a,b,c,d), a także występujące u bakterii fotosyntetycznych – bakteriochlorofile. Chlorofile zbudowane są z pięciopierścieniowej pochodnej porfiryny zwanej feoporfiryną, zawierającej sprzężony układ wiązań podwójnych, zapewniający absorpcję promieniowania świetlnego. W ich skład wchodzą także fitol (dwudziestowęglowy alkohol umożliwiający odpowiednie wbudowanie się chlorofilu w błonę) oraz atom magnezu, odpowiadający za agregację cząsteczek barwnika (ułatwienie przekazu energii wzbudzenia między kolejnymi barwnikami) [15]. Wzory strukturalne chlorofili a i b, wchodzących w skład kompleksów zbierających światło roślin wyższych przedstawiono na rysunku 4.1.
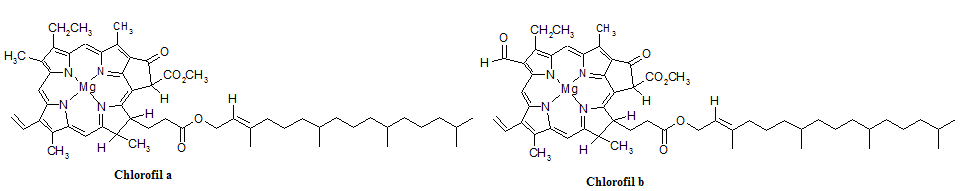
Rys. 4.1. Wzory strukturalne chlorofili a i b
Chlorofil c (c1 i c2) (Rys. 4.2) różni się morfologicznie od pozostałych brakiem fitolu przy węglu C17 [17]. W jego miejscu znajduje się kwas propenowy (akrylowy) w pozycji trans [18].
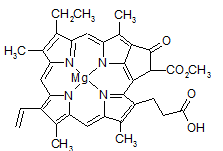
Rys. 4.2. Wzór strukturalny chlorofilu c1
Chlorofile c różnią się od reszty dodatkowo tym, że są magnezofitoporfirynami, a nie magnezochlorynami (zawierającymi całkowicie nienasycony pierścień tetrapirolowy) [18]. Początkowo udało się scharakteryzować jedynie chlorofile c1 i c2, jednak w wyniku zastosowania zaawansowanych technik separacyjnych oraz metod spektroskopowych poznano kolejne pięć rodzajów. Ogólnie przyjmuje się, że chlorofile c1 i c2 pełnią funkcję zbierania światła. Barwniki te są szeroko rozpowszechnione u złoto-brązowych glonów eukariotycznych (okrzemek), gdzie wraz z Chla i karotenoidami tworzą barwniki zbierające światło i wraz z białkami tworzą kompleks antenowy zwany białkowo-chlorofilowo-fukoksantynowym (FCP, ang. fucoxanthin-chlorophyll protein complexes) [18].
Chlorofile nie rozpuszczają się w wodzie, ale możliwa jest ich ekstrakcja rozpuszczalnikami organicznymi. Ponieważ nie absorbują światła z zakresu odpowiadającego światłu zielonemu, ich zabarwienie jest zielone (Chla – niebieskozielone; Chlb – żółtozielone) [14].
Barwniki te poza absorpcją promieniowania pełnią również funkcję polegającą na rozpoczęciu reakcji fotosyntetycznej poprzez przekaz elektronów na specyficzną parę cząsteczek Chla, znajdującą się w centrum reakcji. Proces ten zachodzi z udziałem barwników antenowych (np. karotenoidów). Energia przenoszona jest na kolejne cząsteczki barwników i ostatecznie doprowadzana jest do centrum reakcji fotosyntetycznej (Rys. 4.3) [15].
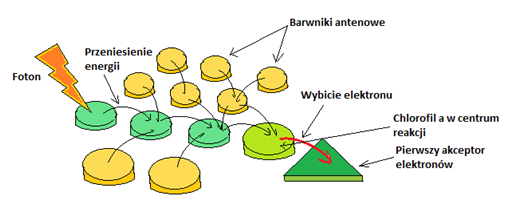
Rys. 4.3. Przekaz energii w błonie tylakoidów z udziałem barwników fotosyntetycznych
Karotenoidy
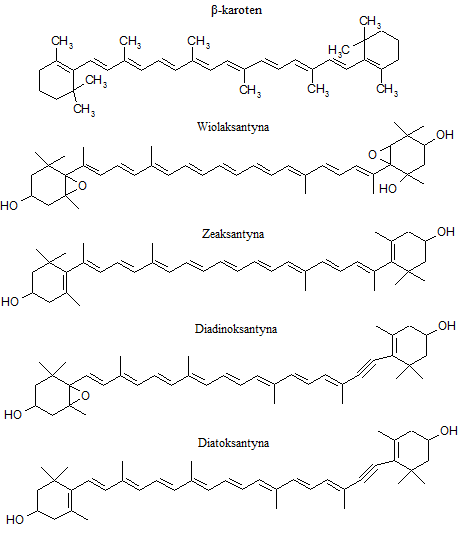 Tworzą podgrupę związków izoprenoidowych zawierającą ponad 700 struktur [19]. Stanowią największą grupę naturalnych barwników. Powszechnie występujące karotenoidy zbudowane są z dwóch pierścieni połączonych łańcuchem węglowodorowym zawierającym sprzężony układ wiązań podwójnych. Dzieli się je na pomarańczowoczerwone karoteny (węglowodory) oraz żółto-pomarańczowe ksantofile (utlenione pochodne karotenów). Do karotenów zalicza się m.in. β-karoten, natomiast do ksantofili np. Ddx, Dtx, Vx, Zx oraz Fx (Rys.4.4. i 4.5.) [19]. Wymienione ksantofile biorą udział w cyklu ksantofilowym i w większym lub mniejszym stopniu występują u wykorzysty- wanych do badań okrzemek.
Tworzą podgrupę związków izoprenoidowych zawierającą ponad 700 struktur [19]. Stanowią największą grupę naturalnych barwników. Powszechnie występujące karotenoidy zbudowane są z dwóch pierścieni połączonych łańcuchem węglowodorowym zawierającym sprzężony układ wiązań podwójnych. Dzieli się je na pomarańczowoczerwone karoteny (węglowodory) oraz żółto-pomarańczowe ksantofile (utlenione pochodne karotenów). Do karotenów zalicza się m.in. β-karoten, natomiast do ksantofili np. Ddx, Dtx, Vx, Zx oraz Fx (Rys.4.4. i 4.5.) [19]. Wymienione ksantofile biorą udział w cyklu ksantofilowym i w większym lub mniejszym stopniu występują u wykorzysty- wanych do badań okrzemek.
Rys. 4.4. Wzory strukturalne wybranych karotenoidów
Główny ksantofil okrzemek to Fx, która odpowiada za ich brunatne zabarwienie. Występuje ona w kompleksie zbierającym światło FCP i absorbuje promieniowanie świetlne w zakresie 500-560 nm [20].
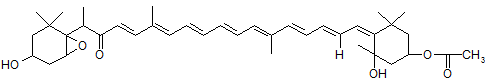
Rys. 4.5. Wzór strukturalny fukoksantyny
Do ważnych ksantofili okrzemek zalicza się także barwniki cyklu diadinoksantynowego, których właściwości fizyczne i chemiczne zamieszczono w tabeli 4.2. Zarówno Ddx jak i Dtx, podobnie jak Fx, wchodzą w skład kompleksu zbierającego światło FCP.
Tabela 4.2. Właściwości fizyczne i chemiczne diadinoksantyny i diatoksantyny [SCOR WG 78 data]
-
Diadinoksantyna Diatoksantyna Wzór sumaryczny C40H54O3 C40H54O2 Masa cząsteczkowa (g/mol) 582,86 566,87 Molowy współczynnik ekstynkcji (l mol-1cm-1) 130∙103 (dla 475 nm w acetonie)
119∙103 (dla 425 nm w acetonie)
Barwa żółta blado pomarańczowa
Karotenoidy podobnie jak chlorofile nie rozpuszczają się w wodzie. Pełnią funkcję anten fotosyntetycznych, tzn. przekazują energię wzbudzenia na cząsteczkę chlorofilu w centrum reakcji, co wiąże się z różnicą w zakresie absorpcji światła przez karotenoidy (400 – 550 nm). Barwniki te ponadto chronią aparat fotosyntetyczny, a szczególnie nienasycone kwasy tłuszczowe lipidów chloroplastowych przed szkodliwym działaniem tlenu w stanie singletowym. Generowany jest on pod wpływem tripletowego stanu chlorofili, który może pojawić się przy intensywnym świetle. Karotenoidy dezaktywują wzbudzony chlorofil, redukując także tworzenie się tlenu singletowego. Ich działanie w tym względzie polega na regulacji przepływu energii między chlorofilem w stanie singletowym i trypletowym [15].
komentarze
Copyright © 2008-2010 EPrace oraz autorzy prac.