
Budowa fotoukładów i ich anten energetycznych roślin wyższych
Pomimo szerokiej różnorodności organizmów fotosyntetycznych na Ziemi, jedynie dwa rodzaje fotoukładów występują w przyrodzie. Znajdują się one zarówno w błonach fotosyntetycznych roślin wyższych, jak i glonów, a nawet bakterii fotosyntetyzujących. Pierwszy z nich to fotoukład I (PS I), a drugi to fotoukład II (PS II). Oba różnią się między sobą budową, składem barwnikowym, lipidowym, białkowym oraz właściwościami spektralnymi (Rys. 4.6.). Zarówno w przypadku PS I jak i PS II centrum reakcji stanowi para Chla, jednak występują różnice we właściwościach spektralnych. Wiąże się to z różnym otoczeniem białkowo-lipidowym w błonie. Dla PS I maksimum absorpcji występuje przy 700 nm (P-700), natomiast dla PS II przy 680 nm (P-680). Fotoukładom towarzyszą anteny energetyczne: bezpośrednio z nimi połączone oraz anteny peryferyjne. W błonie tylakoidu lądowych roślin zielonych znajduje się również duża antena – kompleks zbierający energię świetlną (LHC, ang. Light harvesting complex) [15].
PS II zawiera około 250 – 300 cząsteczek chlorofilu. Składa się z części rdzeniowej oraz antenowej (anteny bezpośrednie – CP43, CP47; anteny peryferyjne). W skład rdzenia wchodzą polipeptydy D1 i D2, cztery cząsteczki Chla, 2 cząsteczki feofityny, 2 cząsteczki β-karotenu oraz cytochrom b559. Polipeptydy D1 i D2, tworzące heterodimer D1/D2, wiążą centrum reakcji fotochemicznej P-680 oraz feofitynę [15].
Główną część anten peryferyjnych stanowi kompleks LHC II (ang. Light harvesting chlorophyll a/b protein complex associated with photosystem II), zawierający około 50% chlorofilu całej rośliny. Uważa się, iż jest to białko występujące w największej ilości w błonach tylakoidowych. LHC II zbudowany jest z 232 aminokwasów, 12 – 15 cząsteczek Chla i Chlb, z jednej cząsteczki neoksantyny, Vx i z jednej lub dwóch cząsteczek luteiny. Poza barwnikami i białkami w kompleksach tych zidentyfikowano jeszcze: fosfatydyloglicerol (PG) i digalaktozylodiacyloglicerol (DGDG) [15]. Jest to jedyny kompleks antenowy, dla którego udało się otrzymać trójwymiarowy model strukturalny za pomocą krystalografii elektronowej. Pozwoliło to ustalić zawartość Chla i Chlb, pozycje 12 cząsteczek chlorofilu (7 cząsteczek Chla i 5 cząsteczek Chlb) oraz wszystkich cząsteczek karotenoidów [21, 22].
Główny kompleks zbierający światło LHC II stanowi 30 % całkowitej ilości białka zawartego w membranach chloroplastów roślin wyższych. Spełnia on cztery główne funkcje w procesie fotosyntezy u roślin:
pobiera energię słoneczną i przenosi ją do centrów reakcji w PS I i PS II;
odpowiada za rozmieszczenie wzbudzonej energii pomiędzy oboma fotoukładami;
odpowiada za organizację roślin poprzez utrzymywanie ścisłego ułożenia membran tylakoidów w granach chloroplastów;
przyczynia się do ochrony układu fotosyntetycznego przez wygaszanie reaktywnych produktów ubocznych, takich jak tlen w stanie singletowym i wolne rodniki [23].
PS I zbudowany jest inaczej niż PS II. Część rdzeniowa zawiera centrum reakcji (dimer PSI-A i PSI-B), łańcuch transportu elektronów, około 100 cząsteczek Chla oraz około 20 cząsteczek β-karotenu. Do dimeru przyłączone są kompleksy zbierające światło LHC I, w skład których wchodzi około 70 cząsteczek Chla i Chlb oraz około 15 cząsteczek karotenoidów. LHC I spełniają funkcję związaną z absorpcją światła i przekazaniem energii wzbudzenia do części rdzeniowej, gdzie może być dalej wykorzystana w centrum reakcji [15]. Struktura ta zbudowana jest z czterech kompleksów Lhca, których ważną cechą jest obecność form czerwonych (ang. red forms), tzn. chlorofili wykazujących silne przesunięcie batochromowe oraz szeroki zakres absorpcji i fluorescencji. Dowiedziono, że czerwone formy LHC I absorbują światło w zakresie 705 – 712 nm. Występują one u roślin, glonów i bakterii, jednak ich funkcje nadal nie są do końca poznane [24].
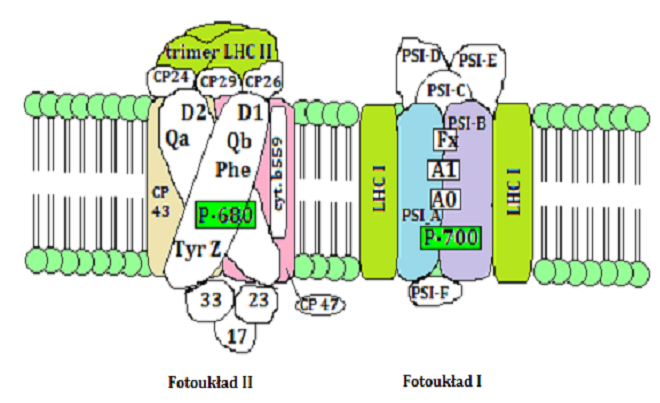
Rys. 4.6. Budowa fotoukładów roślin wyższych [15]
komentarze
Copyright © 2008-2010 EPrace oraz autorzy prac.